Biometria
Variancia,
szórás, t-próba, F-próba
'A' és 'B' populációból vett minták
átlagai és szórásai különböznek-e?
|
|
A populáció |
B populáció |
|
1 |
131.2 |
110.9 |
|
2 |
123.4 |
110.6 |
|
3 |
125.3 |
104.7 |
|
4 |
138.0 |
102.3 |
|
5 |
134.4 |
105.1 |
|
6 |
129.4 |
109.7 |
|
7 |
129.0 |
104.7 |
|
8 |
139.3 |
101.8 |
|
9 |
139.1 |
107.8 |
|
10 |
121.3 |
112.2 |
|
11 |
134.9 |
108.2 |
|
12 |
140.9 |
101.3 |
|
13 |
131.4 |
112.8 |
|
14 |
127.7 |
108.8 |
|
15 |
133.0 |
111.4 |
|
16 |
143.1 |
100.7 |
|
17 |
127.7 |
114.2 |
|
18 |
116.7 |
113.4 |
|
19 |
131.1 |
101.4 |
|
20 |
149.0 |
100.1 |
|
 : : |
132.3 |
107.1 |
|
åx |
2645.9 |
2142.1 |
|
åx2: |
351197.3 |
229860.1 |
|
s2: |
61.6 |
22.6 |
|
 |
1.8 |
1.1 |
s2 = [åx2
– (åx)2/n]/(n-1)
= (351197.3 - 2645.92 /20) = 61.6
= 
 = √61.6/20
= 1.8
= √61.6/20
= 1.8
A két minta átlaga
eltér-e egymástól?
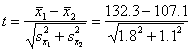 =
25.2/2.109 = 11.9 nagyobb mint a melléklet
t-táblázatban a legközelebbi szabadságfokhoz (40) tartozó P5%
értéke, 2.02 - a két átlag szignifikánsan
különbözik egymástól.
=
25.2/2.109 = 11.9 nagyobb mint a melléklet
t-táblázatban a legközelebbi szabadságfokhoz (40) tartozó P5%
értéke, 2.02 - a két átlag szignifikánsan
különbözik egymástól.
A két minta szórása eltér-e egymástól?
F =  = 2.72 nagyobb mint a melléklet F-táblázatában
a legközelebbi FG[20,20] 2.12 - a két szórás
szignifikánsan különbözik egymástól.
= 2.72 nagyobb mint a melléklet F-táblázatában
a legközelebbi FG[20,20] 2.12 - a két szórás
szignifikánsan különbözik egymástól.
Kovariancia, korreláció, regresszió
Ugyanazon a mintán mért két
tulajdonság (X és Y) között van-e statisztikailag is igazolható összefüggés?
|
n |
X tulajdonság |
Y tulajdonság |
|
1 |
60 |
91 |
|
2 |
73 |
98 |
|
3 |
79 |
98 |
|
4 |
79 |
105 |
|
5 |
82 |
108 |
|
6 |
84 |
104 |
|
7 |
85 |
104 |
|
8 |
88 |
102 |
|
9 |
88 |
99 |
|
10 |
90 |
103 |
|
11 |
95 |
102 |
|
12 |
95 |
108 |
|
13 |
96 |
96 |
|
14 |
98 |
98 |
|
15 |
99 |
111 |
|
16 |
101 |
104 |
|
: |
87 |
102 |
|
åx |
1392 |
1631 |
|
åx2: |
122876 |
166649 |
|
SQ |
1772 |
389 |
|
s2= SQ/FG |
118 |
26 |
SQ = eltérésnégyzetösszeg = [åx2
– (åx)2/n]
, SQx = 122876 – 121104 = 1772
SP
= szorzatösszeg =
åxy
– (åx
*
åy)/n
= 142304 – 141897 = 407
Kovariancia = Covxy = SP/FG = 407/(16-1) = 27.1
Regressziós együttható = bxy = SP/SQx =
407/1772 = 0.23
Regressziós együttható hibaszórása = sb = 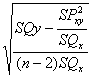 =
0.109
=
0.109
b
regressziós együttható megbizhatóan eltér-e 0-tól?
b/sb
= 2.11 kisebb mint n-2=14 szabadságfokhoz tartozó
táblázati kritikus t-érték -
a két
tulajdonság között nincs statisztikailag is igazolható összefüggés.
Korrelációs együttható =  =
0.48. A korrelációs együttható kritikus értékeit táblázatból (pld. Sváb)
kaphatjuk meg.
=
0.48. A korrelációs együttható kritikus értékeit táblázatból (pld. Sváb)
kaphatjuk meg.
Az átlag komponensei
1.) A búza
kalászhossz öröklődésében milyen génhatás valószínűsíthető a szülők és F1-ük
átlagai alapján:
|
|
n |
átlag (mm) |
s2 |
|
P1
|
10 |
105 |
190 |
|
P2
|
10 |
85 |
160 |
|
F1 |
10 |
116 |
170 |
|
Mivel 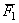 felülmúlja
a nagyobb értékű szülő átlagát is, a feltételezett génhatás típus az
overdominancia. Hipotézisünk tesztelésére ellenőrizzük P1 és
F1 nemzedékátlagok különbségének megbízhatóságát kétmintás
t-próbával :
felülmúlja
a nagyobb értékű szülő átlagát is, a feltételezett génhatás típus az
overdominancia. Hipotézisünk tesztelésére ellenőrizzük P1 és
F1 nemzedékátlagok különbségének megbízhatóságát kétmintás
t-próbával :
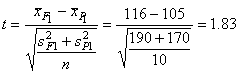
P
= 0.05
valószínűségi-szinten, 2(n-1) =18 szabadság-fokhoz tartozó
táblázati kritikus t-érték 2.1, ami nagyobb a számitott t-nél,
így megállapíthatjuk, hogy a két átlag különbsége nem szignifikáns, azt a
véletlen is okozhatta. Ebből következik, hogy az overdominancia sem nyert
igazolást. Mivel P1 és F1 átlaga
hibahatáron belül megegyező, csak a teljes dominancia fogadható el.
2.) Határozzuk meg két virágzási időben különböző szülő (P1
= 60 nap, P2 = 80 nap) keresztezéséből származó F2
növények fenotípusos értékeit, ha a virágzási időt a szülők különbségének
intervallumában 5 gén (A,B,C,D,E) kontrollálja, és da=db=dc=dd=de,
továbbá -da=-db=-dc=-dd=-de=0,
és ha=hb=hc=hd=he
(lásd a táblázatot).
A két
szülő között 20 nap virágzásbeli különbség van, amit 5 gén, azaz 10 allél
szabályoz, így egy-egy fokozó hatású allél 2 nap tenyészidő növekedést
eredményez (da=db=dc=dd=de=2).
Azon növények, amelyek egyetlen fokozó hatású alléllel sem rendelkeznek (10.
számú növény), a P2 szülővel megegyező fenotípusúak (60 nap).
Mivel a (-) allélek nem járulnak hozzá a fenotípushoz, minden (-/-) lókusz
figyelmen kívül hagyható. Ha nincsenek heterozigóta lókuszok, minden (+/+)
allélpár hatása tisztán érvényesül, 4-4 nappal hosszabbítja meg a tenyészidőt:
például a 3.számú növény esetében 60+2*4=68, mindhárom oszlopban. A heterozigóta
lókuszok dominancia eltéréseinek nagysága mindig a dominancia mértékétől függ.
Ha nincs dominancia (h=0), a heterozigóta lókuszok a (+) allél értékével
(2 nap) járulnak hozzá a fenotípusos értékhez, h=0.5d részleges
dominanciánál 1 nappal, míg h=1.5d overdomianciánál 3 nappal növelik a
tenyészidő hosszat.
|
|
F2
Növények |
Génhatások |
|
Sorsz. |
Genotípus |
|
A |
B |
C |
D |
E |
h
=0 |
h
= 0.5d |
h
= 1.5d |
|
1 |
+/+ |
+/+ |
+/+ |
+/+ |
+/+ |
80 |
80 |
80 |
|
2 |
+/- |
+/+ |
-/- |
-/+ |
-/- |
68 |
66 |
70 |
|
3 |
-/- |
-/- |
+/+ |
-/- |
+/+ |
68 |
68 |
68 |
|
4 |
+/- |
+/- |
+/- |
+/- |
+/- |
70 |
65 |
75 |
|
5 |
+/+ |
-/+ |
+/- |
-/- |
+/+ |
72 |
70 |
74 |
|
6 |
-/+ |
+/- |
+/+ |
-/- |
+/+ |
72 |
70 |
74 |
|
7 |
+/+ |
-/- |
+/+ |
+/- |
-/- |
70 |
69 |
71 |
|
8 |
-/+ |
+/+ |
+/+ |
+/- |
+/+ |
76 |
74 |
78 |
|
9 |
+/- |
+/- |
-/- |
-/- |
+/+ |
68 |
66 |
70 |
|
10 |
-/- |
-/- |
-/- |
-/- |
-/- |
60 |
60 |
60 |
Az F2
növények fenotípusának kiszámítása |
Példaként az 5.számú növény fenotípusos értékének kiszámítása: rendelkezik 2
(+/+) lókusszal, (amelyek 4-4 nappal hosszabbítják meg a tenyészidőt) és 2
heterozigóta lókusszal. h=0 esetén 60+(2*4)+(2*2)=72, h=0.5d
esetén 60+(2*4)+(2*1)=70, és h=1.5d esetén 60+(2*4)+(2*3)=74.
3.) Két eltérő magasságú búzafajta (P1 = 100 cm,
P2 = 80 cm) keresztezéséből kapott F1 magassága
95 cm. A magasabb szülővel (P1 ) három alkalommal
visszakeresztezést végzünk. Milyen magasak lesznek a visszakeresztezett
nemzedékek?
Először határozzuk meg az átlag genetikai komponenseit:
m = (100+80)/2 = 90
[d] = (100-80)/2 = 10
[h] = 95-90 = 5
A három nemzedék átlaga:
 m
+ ½[d] + ½[h] = 90 + (10/2) + (5/2) = 97.5
m
+ ½[d] + ½[h] = 90 + (10/2) + (5/2) = 97.5
 =
90 + {1-(½)2}* 10 + (½)2 * 5 = 98.75
=
90 + {1-(½)2}* 10 + (½)2 * 5 = 98.75
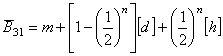 =
90 + {1-(½)3}* 10 + (½)3 * 5 = 99.375
=
90 + {1-(½)3}* 10 + (½)3 * 5 = 99.375
4.) Igaz-e a feltételezés, hogy a táblázatban
szereplő átlagok alapján a példában szereplő tulajdonság egyszerű
additiv-domináns módon öröklődik?
|
|
|
n |
átlag |
s2 |
|
P1
|
100 |
64 |
57,7 |
|
P2 |
100 |
58 |
64,7 |
|
F1 |
100 |
80 |
50,8 |
|
B1 |
100 |
76 |
80,0 |
|
A rendelkezésre álló nemzedékekből az A típusú
lépték próba végezhető el:
A = 2 -
-
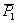 -
-
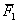 =
2*76 – 64 – 80 = 8
=
2*76 – 64 – 80 = 8
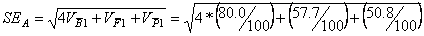 =
6.54
=
6.54
A t-próba szabadságfoka: 3*99=297
t[297] = (8-0)/6.54 = 1.22
Mivel a t-próba nem szignifikáns - a táblázati t-érték (1.96) nagyobb a
számítottnál – így igazoltnak tekinthető az a feltételezés, hogy a tulajdonság
öröklődésében csak a gének additív- és dominancia hatásai játszanak szerepet.
5.) A táblázatban szereplő hat
nemzedék átlagaiból végzett lépték próbák szerint az additív-domináns modell nem
volt illeszthető. Többféle skálát kipróbálva, a lépték próbák a transzformált
adatokkal is szignifikánsak maradtak. Nagy a valószínűsége annak, hogy
nem-alléles gén kölcsönhatással állunk szemben. Számítsuk ki a fő- és
kölcsönhatás paramétereket, és következtessünk a digénes episztázis típusára.
|
|
Nemzedék |
n |
átlag |
átlag varianciája |
|
P1
|
20 |
64.82 |
0.107 |
|
P2 |
20 |
53.07 |
0.365 |
|
F1 |
10 |
55.10 |
0.360 |
|
F2 |
100 |
58.83 |
0.182 |
|
B1 |
10 |
63.15 |
1.822 |
|
B2 |
10 |
59.55 |
0.902 |
|
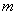 =
½+
½
=
½+
½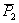 +
4
+
4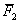 -
2-
2
-
2-
2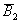 = ½(64.82)+½(53.07)+4(58.83)-2(63.1)-2(59.55) =
48.89
= ½(64.82)+½(53.07)+4(58.83)-2(63.1)-2(59.55) =
48.89
SEm hibaszórás számítása:
SEm = 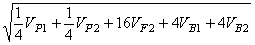 =
=  =
3.733
=
3.733
A becsült paraméter 0-tól való eltérésének megbízhatóságát (a becsült érték
szignifikanciáját) egymintás t-próbával ellenőrizzük:
t[155] = (48.89-0)/3.733 = 13.09 nagyobb mint a táblázati
kritikus t-érték, azaz a becsült
paraméter szignifikáns. A biometriában a becsült
paraméterek szignifikanciáját, az elért valószínűségi szintet a paraméter után,
jobb felső indexbe helyezett csillagok (*) számával szokás jelölni. Megegyezés
szerint * = P0.05, ** = P0.01, *** = P0.001, ha
nincs egyetlen csillag sem, akkor a becsült paraméter nem szignifikáns.
Ezt alkalmazva m = 48.89***, ami azt jelenti, hogy a becsült
m homozigóta közép legalább P=0.001 valószínűségi szinten szignifikáns.
[d] = ½-
½ =
½(64.82)-½(53.07) = 5.87***
[h] = 6+
6-
8- -
1½-
1½=
6(63.15)+6(59.55)-8(58.83)-55.10-1.5(64.82)-1.5(53.07) = 33.57***
[i] = 2+
2-
4= 2(63.15)+2(59.55)-4(58.83) = 10.06***
[j] = 2- -
2+ =
2(63.15)-64.82-2(59.55)+53.07 = -4.55
[l] = + +
2+
4-
4-
4= 64.82+53.07+2(55.10)+4(58.83)-4(63.15-4(59.55) = -27.36
A becsült
paraméterek alapján levonható következtetések: a tulajdonság öröklődésében a
poligének additív- és dominancia hatásai egyaránt jelentősek.
Mivel [h]
előjele pozitív, a lókuszok döntő hányadában a fokozó hatású allélek a
dominánsak. A digénes episztázis paraméterei közül csak az [i]
szignifikáns, azaz csak a gének additív hatásai közti kölcsönhatások
számottevőek. [h] és [l] előjelei különbözőek, a kimutatott
nem-alléles gén kölcsönhatás nagy valószínűséggel komplementer típusú.
Effektiv
faktorok száma
1.) Két homozigóta vonal keresztezéséből származó F2-ben
négy poligén hasad. A populáció hány százaléka hordoz 3 fokozó hatású allélt?
A hasadó gének számát k-val, a fokozó hatású
allélek számát j-vel jelölve:
P3
=  *
*
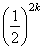 =
=
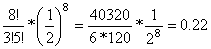 ,
tehát a populáció 22 százaléka hordoz 3 fokozó hatású allélt.
,
tehát a populáció 22 százaléka hordoz 3 fokozó hatású allélt.
2.) Egy F2 populációban a növénymagasság alábbi
megoszlását kaptuk, a fenotípusos megoszlás alapján hány gén hasadása
valószínűsíthető:
|
cm |
növényszám |
|
<50 |
3 |
|
51-55 |
19 |
|
56-60 |
89 |
|
61-65 |
210 |
|
66-70 |
429 |
|
71-75 |
492 |
|
76-80 |
413 |
|
81-85 |
252 |
|
86-90 |
101 |
|
91-95 |
11 |
|
>95 |
4 |
|
összesen: |
2023 |
|
a hasadó génszám és a fenotípus kategóriák száma közötti
összefüggés 2n+1 = x, ahol n a gének és x a
kategóriák száma.
A
példában a kategóriák száma 11, tehát 2n+1 = 11, amiből n=5, azaz
valószínűleg 5 gén hasad.
A
feltételezést ellenőrizzük le statisztikai próbával is!
Ehhez
először meghatározzuk az 5 gén hasadása esetére az elméleti gyakoriságokat. A
táblázat harmadik oszlopában szereplő gyakoriságokat a összefüggéssel
számítottuk. Ezzel megkaptuk az 5 génes hasadás megoszlását a minimális
populációméretre (1024), amit a valós populációméretre (2023) kell átszámítani
(a valós és az elméleti populációméret hányadosával történő szorzással). Az
eredményt a táblázat 4. oszlopa tartalmazza. Továbbiakban a két megoszlás
egyezőségét ellenőrizzük χ2 próbával. Ehhez a két megoszlás
különbségét négyzetre emeljük, osztjuk az elméleti gyakorisággal, és a kapott
eltérésnégyzeteket összegezzük. A számított χ2 összeget (12.9)
hasonlítjuk 11-2=9 szabadság-foknál a kritikus táblázati értékhez. Mivel
számított χ2 értékünk kisebb mint a P=0.05 valószínűségi szinthez
tartozó kritikus érték (16.92), tehát a két megoszlás nem tér el egymástól
statisztikailag igazolhatóan, azaz elfogadjuk a feltételezett 5 génes hasadást.
|
cm |
Gyakoriság |
χ2 |
|
megfigyelt |
elméleti 1 |
elméleti 2 |
|
|
<50 |
3 |
1 |
2 |
0.5 |
|
51-55 |
19 |
10 |
20 |
0.1 |
|
56-60 |
89 |
45 |
90 |
0.0 |
|
61-65 |
210 |
120 |
240 |
3.8 |
|
66-70 |
429 |
210 |
420 |
0.2 |
|
71-75 |
492 |
252 |
504 |
0.3 |
|
76-80 |
413 |
210 |
420 |
0.1 |
|
81-85 |
252 |
120 |
240 |
0.6 |
|
86-90 |
101 |
45 |
90 |
1.3 |
|
91-95 |
11 |
10 |
20 |
4.1 |
|
>95 |
4 |
1 |
2 |
2.0 |
|
összeg: |
2023 |
1024 |
2048 |
12.9 |
|
ÖRÖKÖLHETŐSÉG
1.) Becslése varianciakomponensekből
a.) Véletlen Blokk Elrendezésű kísérlet egy
környezetben, 2 ismétléssel
egy keresztezésből származó 48 db homozigóta vonal
|
Forrás |
FG |
SQ |
MQ |
F |
Szign. |
|
Összes |
95 |
1810.429 |
19.0572 |
|
|
|
Genotípus |
47 |
1388.432 |
29.5411 |
3.4 |
***
|
|
Ismétlés |
1 |
12.9084 |
12.9084 |
1.5 |
n.s. |
|
Hiba |
47 |
409.0893 |
8.704 |
|
|
|
σ2e = MQhiba =
8.70
σ2g = (MQgenotípus
– MQhiba)/ismétlésszám = 10.41
σ2p = σ2e
+ σ2g = 19.11
h2 = σ2g/ σ2p
= 0,54
b.) Ugyanaz mint (a) alatt, de két évben
megismételve
|
Forrás |
FG |
SQ |
MQ |
F |
|
|
Összes |
191 |
9148,13 |
|
47,89 |
|
|
|
Genotípus (G) |
47 |
1452,21 |
M1 |
30,89 |
1,7 |
* |
|
Környezet (K) |
1 |
5440,02 |
|
5440,02 |
302,2 |
*** |
|
Ismétlés/K |
2 |
314,28 |
|
157,14 |
8,7 |
*** |
|
G x K kölcsönh. |
47 |
846,07 |
M2 |
18,00 |
1,5 |
* |
|
Hiba |
94 |
1095,53 |
M3 |
11,65 |
|
|
|
ismétlések száma: r, környezetek száma: t
σ2e = M3 = 11,65
σ2ge = (M2-M3)/r
= 3.17
σ2g = (M1-M2)/rt
= 3,22
σ2p = σ2e
+ σ2ge + σ2g = 18,04
h2 = σ2g/ σ2p
= 0,17
Példa diallél keresztezés
kiértékelésére
Hipotézis:
¨
diploid hasadás
¨
nincsenek a reciprokok között különbségek
¨
a nem-allél gének egymástól függetlenül hatnak
¨
nincs többszörös allélia
¨
a szülők homozigóták
¨
a gének egymástól függetlenül oszlanak meg a szülők között
Þ
Alapadatok véletlen blokk elrendezésű kisérletben
pl: 5 szülő + 10 kombináció, 4 ismétlésben
Alapadatok
variancia-analizise:
|
Forrás |
FG |
SQ |
MQ |
F |
|
Kezelés |
14 |
539.58 |
38.54 |
76.14 *** |
|
Blokk |
3 |
1.8 |
0.60 |
1.19 |
|
Hiba |
42 |
21.25 |
0.50 |
|
Következtetés: a kezelések között statisztikailag
igazolható különbségek vannak, tehát folytatható az elemzés
Þ
Diallél-táblázat, sorösszegek, sorátlagok, (Vr)
és (Wr) számitása
|
Szülők |
P1 |
P2 |
P3 |
P4 |
P5 |
|
P1 |
7.67 |
8.37 |
10.47 |
9.6 |
12.67 |
|
P2 |
|
8.27 |
9.7 |
9.95 |
14.17 |
|
P3 |
|
|
11.65 |
11.15 |
14.16 |
|
P4 |
|
|
|
12.37 |
15.42 |
|
P5 |
|
|
|
|
19.12 |
|
Sorösszeg |
48.78 |
50.46 |
57.14 |
58.49 |
75.55 |
|
Sorátlag |
9.75 |
10.09 |
11.42 |
11.69 |
15.11 |
|
Wr |
8.66 |
10.84 |
7.48 |
10.63 |
10.76 |
|
Vr |
3.82 |
5.77 |
2.88 |
5.51 |
5.97 |
|
W'r |
4.00 |
5.07 |
3.48 |
4.93 |
5.04 |
|
Wr+Vr |
12.84 |
16.61 |
10.37 |
16.15 |
16.74 |
|
Wr-Vr |
4.83 |
5.07 |
4.59 |
5.12 |
4.79 |
a 3. sor varianciája = [10.472+9.72+11.652+11.152+..
..14.162-(57.142/5)]/4
a 3 sor kovarianciája =
[(10.47*7.67)+(9.7*8.27)+(11.652)+(11.15*12.37)+..
..(14.16*19.12)-((57.14*59.08)/5)]/4
W'r 3. sorra =
[(10.47*9.75)+(9.7*10.09)+(11.65*11.42)+..
..(11.15*11.69)+(14.16*15.11)-((57.14*58.08)/5)]/4
Þ
A modell megfelelőségének ellenőrzése:
¨
Wr/Vr regressziós egyenes
bWr,Vr = 1.112 SEb = 0.063
a 0-tól való eltérés próbája: (1.112-0)/0.063 = 17.7
szignifikáns, mert a táblázati t5% FG (p-2) : 3-nál 3.18
Következtetés: a sorok varianciái és kovarianciái között
igazolt összefüggés van
1-től való eltérés próbája: (1.112-1)/0.063=1.77 nem
szignifikáns, mert a táblázati 't' FG 3 nagyobb
Következtetés: a diallélban nincsenek kimutatható torzitó
hatású nem-alléles kölcsönhatások
¨
(Wr-Vr) különbségek homogenitása
(Wr-Vr) különbségek ismétlésenként
|
Sorok
|
R1 |
R2 |
R3 |
R4 |
|
P1 |
9.13 |
14.84 |
23.66 |
8.22 |
|
P2 |
19.64 |
20.67 |
17.09 |
10.82 |
|
P3 |
10.82 |
8.94 |
8.85 |
14.27 |
|
P4 |
13.34 |
19.76 |
16.60 |
16.22 |
|
P5 |
17.96 |
17.09 |
12.71 |
22.63 |
A (Wr-Vr) különbségek szórásnégyzet-elemzése:
|
Forrás |
FG |
SQ |
MQ |
F |
|
Összes |
19 |
4.74 |
0.24 |
|
|
Sorok között |
4 |
1.88 |
0.47 |
2.48 n.s. |
|
Sorokon belül |
15 |
2.85 |
0.19 |
|
Következtetés: a nem szignifikáns sorok közötti (Wr-Vr) különbség is azt jelzi,
hogy a diallélban nincsenek kimutatható torzitó hatású nem-alléles
kölcsönhatások
Þ
Statisztikák becslése:
V0L0
(Vp) = 20.861
V0L2 (Vr) =
4.501
V1L2 (Vr) =
4.790
W0L02
(Wr) = 9.678
ML2
= 11.61
ML0
= 11.81
Þ
Varianciakomponensek számitása
D = 20.73
± 0.166 ***
F = 2.85
± 0.754 ***
H1 = 4.79
± 1.794 **
H2 = 2.98
± 1.624 n.s.
h2 = 8.37
± 1.098 ***
E = 0.12
± 0.067 n.s.
varianciakomponesek statisztikai próbája:
[ D/SE(D) ] hasonlítva táblázati "t"
értékhez
Þ
Egyéb paraméterek becslése:
¨
dominancia átlagos foka:
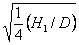 = 0.24
= 0.24
¨
+ és - hatású gének aránya a szülőkben:
H2/4H1
= 0.155
¨
dominancia iránya:
r(Wr+Vr,Yr) = 0.357 n.s.
¨
domináns és recessziv gének aránya a szülőkben:
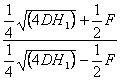 = 1.01
= 1.01
¨
dominanciát mutató lókuszok száma:
h2/H2
= 2.8
Þ
Wr/Vr grafikonból:
¨
szülők dominanciasorrendje
¨
dominancia mértéke
¨
szelekciós előrehaladás becslése